走进仿生超浸润界面专题 | (二)自然超浸润界面
仿生超浸润领域在过去的重要性、现在的影响力和未来的可期、可用、可为,充分说明该领域具有无限潜力和生机活力。只有观大势、明大势,才能应势而动、顺势而为,为此,国家先进功能纤维创新中心以李昶、李明与创新中心赵润博士参与编著的《仿生超浸润界面》为核心,在公众号上展开为期6期的仿生超浸润界面专题科普,希望为读者带来一些科学、文化、前瞻的启发。敬请持续关注公众号最新动态~水是生命之源,生态之基。动植物的生活
仿生超浸润领域在过去的重要性、现在的影响力和未来的可期、可用、可为,充分说明该领域具有无限潜力和生机活力。只有观大势、明大势,才能应势而动、顺势而为,为此,国家先进功能纤维创新中心以李昶、李明与创新中心赵润博士参与编著的《仿生超浸润界面》为核心,在公众号上展开为期6期的仿生超浸润界面专题科普,希望为读者带来一些科学、文化、前瞻的启发。敬请持续关注公众号最新动态~

水是生命之源,生态之基。动植物的生活都离不开水,而自然界中一些生物的界面拥有特殊的操控水分子的能力,并且展现出有趣而又独特的浸润现象、黏附能力及液滴行为。相关内容在前文中有所提及,本章将详细介绍自然界中的超浸润界面及其相关现象。
研究发现,这些生物界面有着特殊的微纳米结构或化学组成分布,导致了界面特殊的浸润性与黏附特性。本章涵盖空气中及水下研究,涉及人类、动物界、植物界、微生物界;并按照浸润/黏附性分类介绍自然界超浸润界面实例,包括超疏水低黏附表面[如图 1(a)、(b)所示]、超疏油/超双疏表面[如图 1(c)所示]、超疏水高黏附表面[如图 1(d)、(e)所示]、超浸润各向异性界面[如图 1(f)、(g)所示]、超亲水表面/水下超疏油界面[例如图 1(i)~(l)所示]。


(a)荷叶上表面 (b)猪笼草 (c)跳虫 (d)玫瑰花瓣 (e)槐叶萍 (f)蝴蝶翅膀


(g)黑麦草叶 (h)松叶 (i)荷叶下表面 (j)蛤壳 (k)海藻 (l)鲀鱼
图 1 动植物超浸润界面及其显微结构展示(代表性界面列举)
超疏水表面上液滴呈现近似球形,可减少固-液接触,通常使表面与水表现出低黏附性质。自然界中生物界面的超疏水低黏附策略包括特殊的微纳米复合结构防止浸润、表面注入润滑物质降低黏附和摩擦、陷入空气以降低液滴运动阻力等。
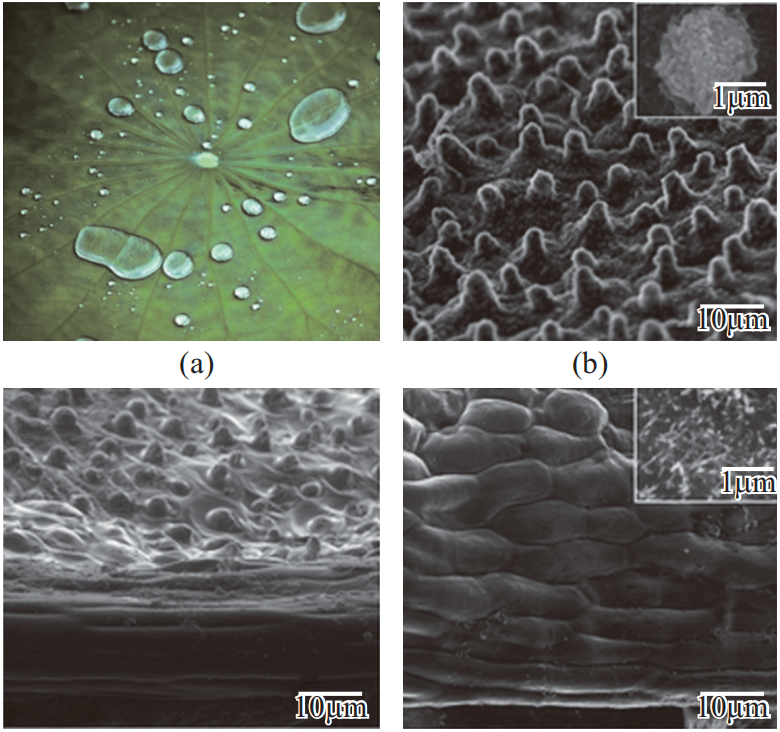
荷叶出淤泥而不染,以自清洁、防污而闻名,是许多文化中纯洁的象征。其不染淤泥背后的科学奥秘在于荷叶表面的超疏水低黏附性质。W. Barthlott 和 C. Neihuis 发现当水滴落在荷叶上时呈近似球体(CA 大约160°),他们认为这与荷叶表层的蜡质和表面微结构有关。液滴很容易从表面滚落,并同时带走黏附的污垢颗粒,即“荷叶效应”。Jiang 等发现超疏水荷叶上不仅有微米结构,还有微纳米多层次的复合结构。如图2(a)~(d)所示,在每个微米级突起结构上可观察到有层次的纳米结构。突起结构减少了水与荷叶表层的接触,利用空气层将水隔开;多尺度结构及表面蜡质可有效防止荷叶表面被浸润,也可预防液滴进入微结构。液滴接触该结构时产生的 TPCL 不稳定,因此容易移动。如图 2(e)所示,根据曲线结果可通过计算给出层次结构和浸润性之间的理论关系,有力证明自清洁性是由于荷叶上多尺度结构和蜡层的协同作用。
(c) (d)
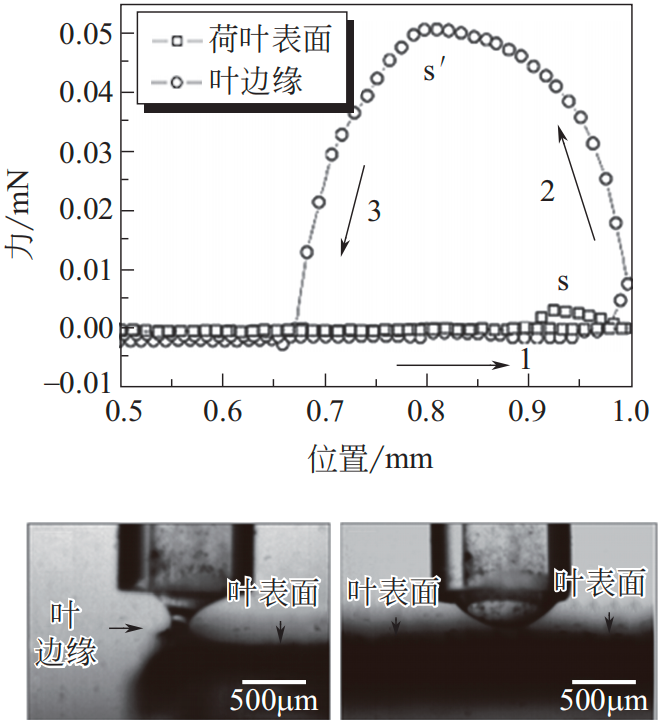
(e)
图 2 荷叶上表面的表面形貌图和超疏水自清洁特性
猪笼草(nepenthes)、瓶子草(pitcher plant)等瓶子草科(Sarraceniaceae)植物,有一个瓶状口袋(叶笼),可消化昆虫等小动物来获取营养。观察发现,当小昆虫接触进入红瓶猪笼草口袋的唇部等红色区域时,会突然掉入口袋。猪笼草具有这种天赋主要是由于其唇口可分泌润滑物质,且与唇口相连的红色区域(被称为“滑移区”,即“slippery zone”)也富含润滑蜡质。
当物体接触其表面时,猪笼草可分泌出润滑物质注入微纳米结构,导致表面油滑,生物或液滴难以保持静止。猪笼草的唇口和滑移区具有不同的结构及浸润性,以满足特定需求。如图 3 所示,唇口表面具有密布的条纹微槽,其边缘具有弓形的倒钩状结构,可阻碍生物撤退;在未充满油滑物质时表现出超亲水性质,水滴会在微槽中定向流动,从结构中被排出,该结构可防止其他液体堆积或倒流入袋;在分泌出大量润滑物质后,该结构可避免润滑物质流失,并且使表面润滑,有助于生物滑入口中。小动物在经过唇口后首先接触到红色“滑移区”,该区域的富含润滑蜡质层,有弯月形微阵列结构,表现为超疏水低黏附特性。
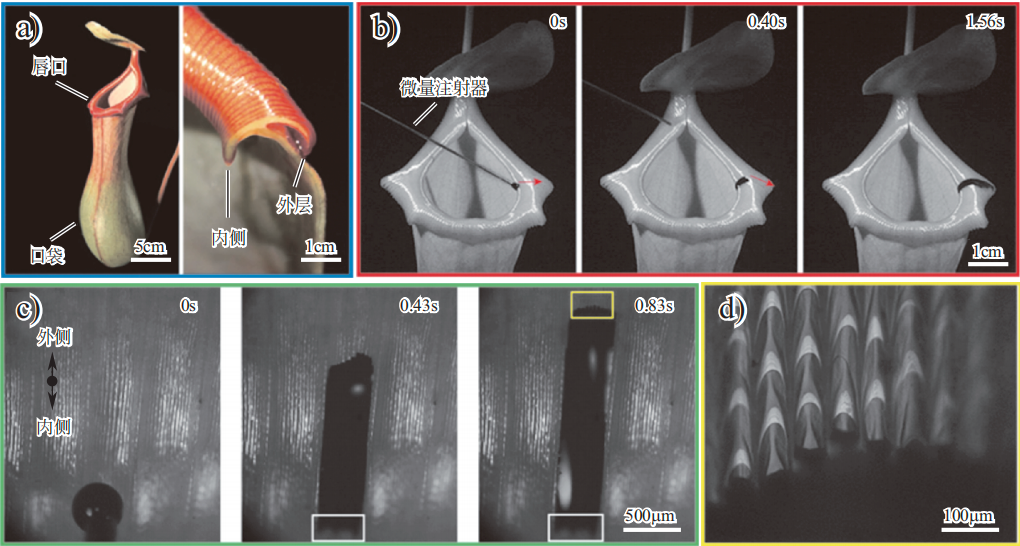

图 3 猪笼草的唇口表面形貌和超浸润油滑特性
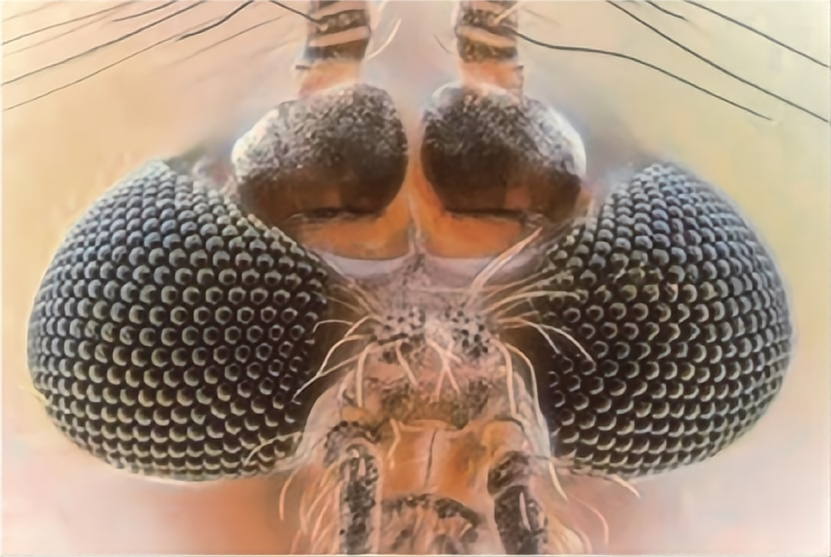
即使在昏暗潮湿的栖息地,蚊子依然具有出色的视力。Jiang 等发现,蚊子复眼具有超疏水防雾功能。该超浸润性主要归功于其特殊的微纳米复合结构,即复眼由密布的六边形小眼组成,其上还有突起结构,如图 4 所示。密布的多级结构可产生疏水毛细作用力,保留微纳米结构中陷入的空气防止液体浸润,即使受到一定压力也可使表面液滴保持 Cassie 态,即展现超疏水低黏附性质。即使在潮湿条件中,微小雾滴也难以在复眼表面凝结。
(a)

(b)

(c)

(d)

(e)

(f)
图 4 蚊子眼睛的表面形貌和超疏水防雾性能
普通的超疏水低黏附表面虽然能够防止被水浸润,但仍可被油类等液滴污染。自然界的生物界面中,还有着不仅能够超疏水还能超疏油的超双疏表面。这些具有超双疏表面的生物通常生活在更为恶劣的脏污环境。超疏油(超双疏)表面要求材料表面能更低,对于材料的化学组成、微纳米结构等要求更苛刻。
由于条件更为苛刻,当前在自然界发现的超双疏表面例子较少,主要包括枯草芽孢杆菌[图 5(a)]、叶蝉[图 5(b)]、各种跳虫[图 5(c)~(f)]等。除具有更为特殊的化学组成外,这些表面的相关微纳米结构具有更多复杂的层次或更精细的纳米形貌。
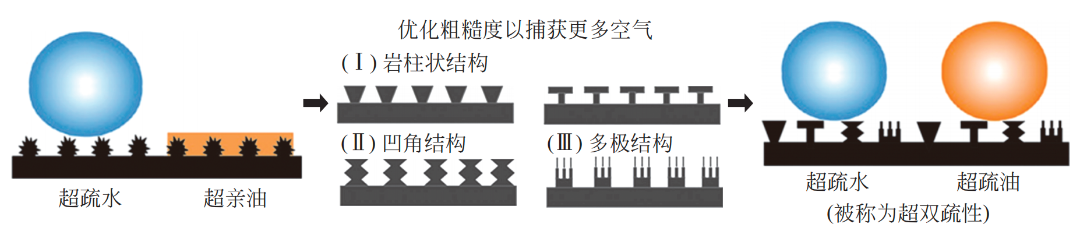
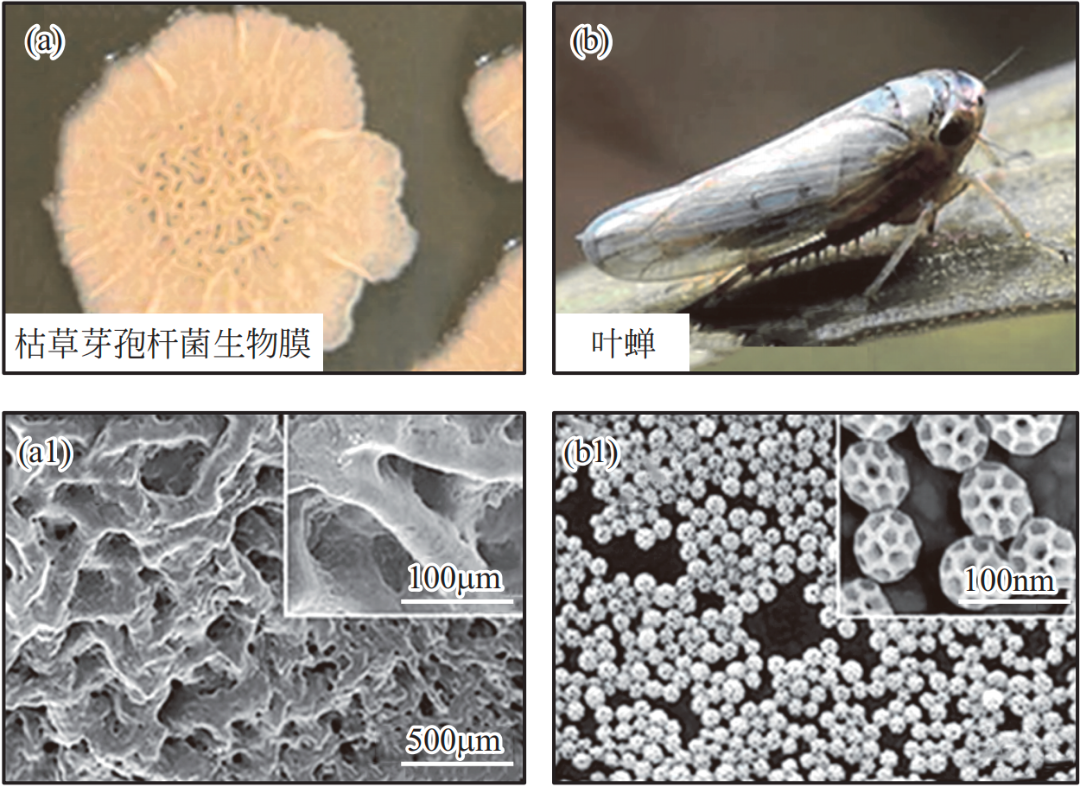



图 5 自然界发现的超双疏表面
枯草芽孢杆菌(Bacillus subtilis)是一种具有很强环境适应性的微生物,在环境恶劣、营养物质缺乏的环境下会进入孢子休眠期,可较长时间生存于高温、酸碱等恶劣环境。Aizenberg 等发现,枯草芽孢杆菌的生物膜具有很强的保护机制,即使处于高浓度酒精等低表面张力液体浓度达到所谓的“致死浓度”的环境中,一定时间内仍可以防止液体渗透到其生物膜结构中。该生物膜表面可保持高度疏液状态(水、酒精的接触角为 115° ~ 150°)。研究认为,其生物膜表面由特殊的蛋白质分子组成,可防止表面被一些液体浸润;此外,如图 5(a)所示,该生物膜表面的微米皱褶、多级凹角结构(reentrant curvatures/structure)也被认为是表面不被润湿的关键。
叶蝉(leafhoppers)常见于农林业作物,繁殖力强,以植物为食并分泌有害物质,可传播植物病毒病。人类往往需要通过增加杀虫剂浓度或剂量以应对叶蝉灾害;长此以往,一些叶蝉甚至会对某种农药产生抗药性,需要更换农药或采用如灯光诱杀等其他手段才可达到灭杀目的。叶蝉能够抵御一些杀虫剂可能与其外皮覆盖物(integuments)的超浸润性有关。Rakitov 和 Gorb发现叶蝉的外皮覆盖物的表面有许多均匀密布的brochosomes结构,如图 5(b)所示。该结构由直径为 200 ~ 700nm的具有蜂窝状内壁的空心蛋白质球组成,不仅可防止自身分泌物残留、黏附在外皮表面,也可防止外来液体浸润。叶蝉外皮覆盖物对水、二碘甲烷、乙二醇等液体都表现出超疏液性、排液性,仅不能有效阻挡乙醇。
实验证明,其表面复杂结构的分形(fractal)粗糙是其超双疏的关键,旨在最大程度减少表面与液体的接触。
跳虫(springtail,collembolan)常生活在腐烂有机物、有害微生物等高度污染的潮湿环境中。跳虫能够抵御恶劣环境的影响,原因在于跳虫皮层的甲壳素(chitin)、蛋白质等化学组成的性质稳定,并且其皮层的特殊结构可阻止有害物质入侵。研究表明,跳虫皮层具有超双疏性质,不仅超疏水,而且可以阻止一些有机液体浸润;将跳虫淹没在水或一些有机液体(如乙醇、油等)中,其皮层表面可形成保护气垫防止浸润,跳虫甚至可以由此呼吸空气,避免窒息。如图 5(c)~(f)所示,不同的跳虫都具有类似的复杂纳米凹角结构,如多边形凹角、蘑菇状结构等,这些凹角结构是形成气垫、防止油滴浸润的关键。
一些种类的玫瑰花瓣具有类似荷叶的超疏水低黏附性质及自清洁能力,然而其他种类则表现出不同的性质。在花园中常常见到,红玫瑰(red rose)的花瓣(petal)上总是沾满晶莹剔透的露珠,这些露珠呈近似球体(超疏水性质)并始终黏附在花瓣表面(高黏附特性)。Feng 等研究表明该种红玫瑰花瓣上 WCA=152.4°,而即使将花瓣倒置,液滴仍然黏附在花瓣上不掉落。这种在界面上表现出超疏水但高黏附的现象被称为花瓣效应(petal effect)。如图 6 所示,与荷叶结构相似的是,界面上密布的微突起阵列是超疏水的原因之一;然而,相较于荷叶的微纳米结构,该微突起尺度较大且表面有明显的纳米褶皱,这使得液滴容易进入微结构的间隙并使 TPCL 维持较稳定的状态,导致较大的接触角滞后,即玫瑰花瓣的高黏附性质。

图 6 红玫瑰花瓣的表面形貌和超疏水高黏附性质
此外,红玫瑰花瓣的鲜艳色彩也与纳米结构导致的结构色有关。自然界还有一些超浸润结构表面同时具有结构色。例如,在夏季,白杨树的树叶会呈现白色。研究表明,白杨树叶表面具有超疏水结构,该结构同时具有高反射性,可保障白杨树叶不被夏季的烈日灼伤。但结构色与浸润性之间是否有联系有待于跨领域研究者进一步探索。例如,其他颜色的玫瑰花瓣表现普通疏水或超疏水低黏附性质;一些青杨树叶仅表现出普通疏水状态(WCA 约为 110°)。
不同的环境下可发现具有不同超浸润特性的生物界面。主要生活在干旱、半干旱区域的花生,其花生叶(peanut leaf)与水生荷叶不同,而与红玫瑰花瓣相似,展现出超疏水高黏附特性[图 7(a)]。Liu 等发现花生叶 WCA > 150°,但液滴在花生叶表面的黏附力可达 70μN;而荷叶具有类似的 WCA,但黏附力却趋近于 0μN。他们研究了花生叶的微纳米结构[图 7(b)~(d)],并认为其超疏水高黏附特性是由于微米尺度下准连续的 TPCL 和纳米尺度下非连续的 TPCL 造成的。更有趣的是,Jiang 等进一步研究表明该超疏水性质在高湿环境下会被破坏,居然可直接转变为超亲水性。当相对环境湿度(relative humidity,RH)> 99% 时,高蒸气压的水分子与微纳米结构之间的作用力可使液滴凝结,并浸没微纳米结构,使表面展现超亲水状态。


图 7 花生叶的表面形貌[(a)~(d)]和超疏水高黏附性质及高湿环境下的超亲水性质(e)
槐叶萍(Salvinia)是一种水生浮萍,由于其表面特殊的微纳米结构、生物化学组成分布及超浸润性,使其能在水面漂浮。研究表明,其表面的微观结构为大约 2mm 高的柱状阵列,每个柱状结构的顶部长有触手,使阵列整体结构形如打蛋器,如图 8 所示;柱状阵列覆盖有疏水蜡质晶体,而顶端触手则无蜡质。微阵列结构及每个柱状结构的表面疏水性导致整个表面具有超疏水性,而顶部的亲水性导致表面展现出对于水的高黏附性。在单一整体结构中具有离散性质的现象也因此被称为“槐叶萍悖论(Salvinia paradox)”。
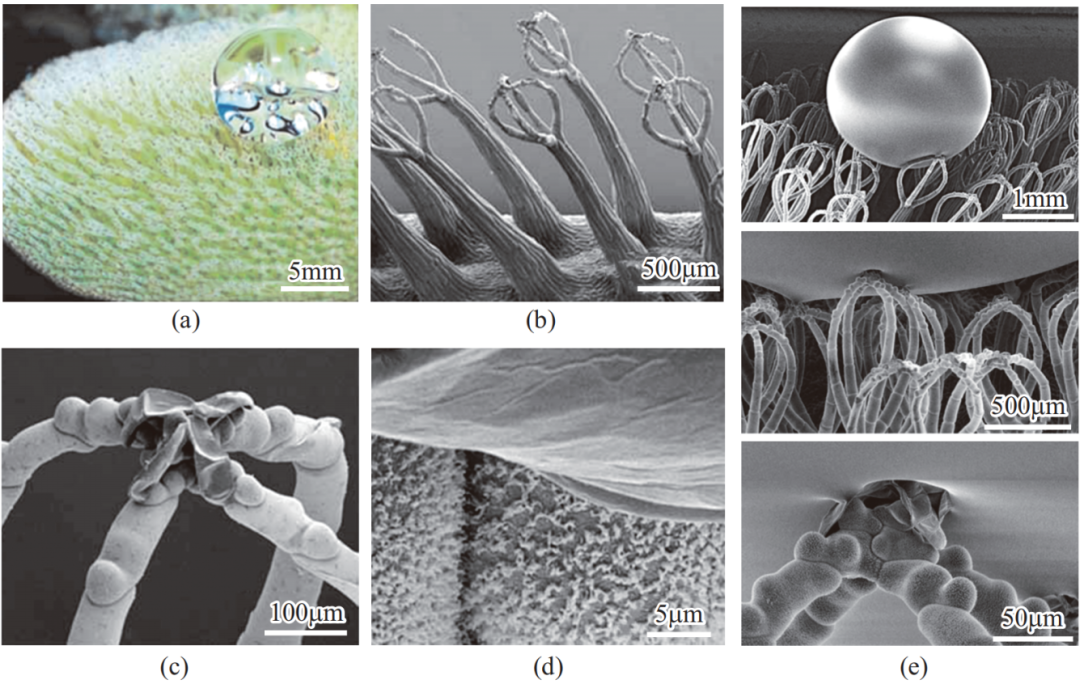
图 8 槐叶萍的表面形貌和超疏水高黏附性质
这种超疏水高黏附的策略可赋予表面更多功能。例如,与同为超疏水高黏附的花生叶相反,即使将槐叶萍放入水中,由于顶部亲水而下层及整体疏水,水层仅保持在微纳米顶部以上而不浸没表面结构,该结构表现出良好的保气性(air-retention)和水下超疏水特性。这种将超疏水材料放入水中后在材料表面仍保有稳定空气层的现象,被进一步称作“槐叶萍效应(Salvinia effect)”。
壁虎因其能在墙壁上自由爬行而不摔落闻名。原因在于,壁虎脚掌具有特殊的纳米纤毛结构,在与固体表面接触时产生的范德华力使其可粘在墙壁,甚至倒悬在天花板上。除众所周知的固 - 固可逆黏合性能外,壁虎脚掌刚毛的特殊结构还表现出超疏水性,并基于相似原理对水具有高黏附性。
Autumn 等发现壁虎刚毛的 WCA 约为 160°。Liu 等进一步研究了壁虎脚掌黏附力随浸润状态的变化,如图 9 所示。壁虎脚掌可调控与接触物体的黏附力,主要归因于接触条件(如距离、压力、接触面积等)的变化、脚掌刚毛接触物体时表面蛋白质的构象转化等。



图 9 壁虎脚掌的表面形貌图和超疏水高黏附性质
一些表面上的微纳米结构或化学组成分布具有取向性,并可产生由粗糙度差或异质组成导致的浸润性/表面能梯度、几何梯度导致的 Laplace 压差等作用力。这些结构及作用力对于液滴运动的影响表现为:液滴在表面的运动具有方向性,或/和液滴朝不同方向运动时的阻力大小存在显著差异。自然界中,各向异性表面十分常见,很多生物都需要有这种操控液滴运动取向的能力以满足特定生存需求。在各向异性表面上,材料整体的浸润性(接触角)仅影响液滴运动的宏观形式,如一般亲水或疏水的各向异性表面上液滴可定向滑动,超疏水各向异性表面上液滴可定向滚动、跳动,具有取向结构的超亲水通道中的液体可定向流动。
下雨时,蝴蝶也可不受雨滴的影响而翩翩起舞。蝴蝶在雨中飞翔的奥秘在于其翅膀有独特的微纳米结构及相应的润湿性能。与荷叶无序的各向同性结构不同,蝴蝶翅膀表面存在有取向的微米鳞片、纳米条纹、纳米尖端的层次结构,如图 10 所示。研究表明,这种微纳米结构可导致超疏水性(WCA 约为 150° ~ 154°),从而排斥液滴。更重要的是,具有方向性的结构导致各向异性黏附,当蝴蝶挥动翅膀时,如果表面有液滴,则液滴会朝向翅膀外侧运动(即图 10 中的 RO 方向,RO 方向即为液滴滚落方向),直至离开表面。如图 10(b)所示,液滴朝向外侧滚离的水滚动角(WSA)为 10°;而在测试液滴沿相反方向的 WSA 时,发现液滴固定在表面上[图 10(c)],即液滴不会向蝴蝶躯干部分滚动。一些理论指出,这种各向异性与蝴蝶翅膀的取向性结构有关,即液滴向外侧运动时 TPCL 不连续,而液滴向内侧运动时,TPCL 近似连续。一些动物的羽毛也存在类似蝴蝶翅膀的取向性微纳米结构,如鹅毛、鸭毛等都展现出各向异性黏附,其上液滴的行为具有方向性,使其能在水上游。

(a)

(b) (c)

(d)

(e)

(f)
图 10 蝴蝶翅膀的表面形貌和超疏水各向异性黏附
水黾以在池塘、沼泽等缓流液面上自由行走而闻名。Bush 等发现水黾主要通过其毛茸茸的足部产生半球形涡流驱动,将动量传递给下面的流体。Jiang 等进一步揭示了水黾能够站在水面上的奥秘在于其足部结构的超疏水作用。如图 11 所示,水黾足部长有取向性针状微结构的刚毛,与水面呈约 20° 的倾斜角;每根刚毛上还有着取向性纳米凹槽结构。

图 11 水黾足部的表面形貌和超疏水各向异性防浸润
这种微纳米结构可以有效地捕获、保留空气,在足部和水之间形成气垫,赋予足部强大的防湿能力(WCA 高达 168°)。与蝴蝶翅膀的原理类似,取向性结构也有助于在恶劣条件下防止浸润。事实上,即使在狂风暴雨或洪流中,水黾也能浮于水面,仅一只足部就足以承载水黾自身 15 倍的质量。
荷叶是植物界中各向同性超疏水现象的典型代表,液滴可在荷叶表面向各个方向自由滚动。而自然界还有一些植物,为满足生存需求,不仅需要表面超疏水低黏附,还需要操控液滴运动的方向。水稻(rice)是人类最重要的粮食作物之一,主要生长在温带、热带等较高温度的多湿环境下。如图 12 所示,水稻叶表面不仅具有微米沟槽结构,还有类似于荷叶表面微纳米结构的纳米突起结构。Jiang 等研究表明,突起沿着叶缘方向近似为一维有序排列,而垂直于叶缘方向则类似于荷叶的随机排列。由于表面的微米沟槽及其纳米结构排列均具有取向性,液滴沿不同方向运动时需克服的能垒存在差异(沿叶缘方向,WSA 为 3° ~ 5°;而垂直于叶缘方向,WSA 为 9° ~ 15°),这导致水稻叶上的液滴更倾向于沿着叶缘方向一维运动,即滚向自身根部或滚离叶子,这是水稻叶操控液滴满足生存需求的前提之一。

图 12 水稻叶的表面形貌和超疏水各向异性
黑麦草(ryegrass)是广泛分布在欧洲、亚洲、非洲等地的禾本科草类植物,常用于高尔夫、足球等运动草坪,也常作为牛、马、羊、兔等放牧用牧草。与水稻叶、蝴蝶翅膀相似,黑麦草的叶子也具有超疏水各向异性黏附性。然而,这种各向异性是由不同的微纳米结构所导致的。如图 13 所示,Zheng 等在黑麦草叶子表面发现了斜角结构——倾斜的微纳米刺状阵列(即“倒刺”),其中倒刺与平面大约呈 25° 角。研究发现,液滴朝向倒刺倾斜方向运动时 WSA 约为 12°,而沿着相反方向运动时,WSA 约为 26°。研究表明,对于倒刺等具有斜角结构的水平表面(指微纳米阵列结构与表面基底呈非 90°、非 0° 或非 180° 夹角,表面本身不斜置),其上水滴沿着斜角微纳米结构的倾斜方向运动时将被加速,逆方向运动将受到阻碍,即表现出各向异性黏附。

图 13 黑麦草的表面形貌和超疏水各向异性黏附
纳米布沙漠(Namib desert)是世界上最古老的干旱沙漠之一。该沙漠常年少雨,生物无法直接被水滋润。白昼炎热干旱,仅在夜间及清晨一段时间内,温度降低(液滴容易凝结),在该沙漠中生存的生物主要靠此时段从空气中汲取水分。纳米布沙漠甲虫(desert beetle,下文简称沙漠甲虫),如图 14 所示,其背部是亲水突起和疏水纹理交错的异质表面,被认为是制备仿生集水材料的重要模型之一。
仿沙漠甲虫背部异质结构表面操控液滴的主要机理是:亲水部分能够从雾气中快速捕捉水分凝结液滴,疏水部分则有助于防止液滴滞留以便输运,亲疏交错结构给液滴传输提供了方向、路径,即液滴被亲水部分捕捉至表面后,由于异质结构产生的浸润性梯度可驱动液滴定向移动。

图 14 沙漠甲虫表面形貌
蜘蛛网(spider web)有着特殊的拓扑结构,材料本身还含有特殊的生物蛋白成分,因优异的力学性能、生物相容性、医药学功用等被诸多领域的科技人员广泛研究,常见于各领域的仿生研究。蜘蛛网亲水可被润湿,Zheng 等通过观察单根蜘蛛丝(spider silk)发现,其被润湿后变成周期性纺锤节纤维,如图 15(a)所示。纺锤节具有锥度,能够产生 Laplace 压差,给纤维上的液滴施加驱动力使其向纺锤节的中心移动,有利于液滴传输合并成大液滴以掉落收集,如图 15(b)所示。此外,蜘蛛丝的纺锤节部分相较于连接纤维段有着更高的轴向粗糙度,纺锤节部分与水的接触角更小,即表面能更高、更亲水,这也有助于驱动液滴移动、合并。

图 15 蜘蛛丝的表面形貌和液滴定向输运
仙人掌(cactus)作为可在沙漠环境中生存的代表性植物,受到了广泛的研究。其界面液滴输运甚至可实现集水的奥秘主要在于仙人掌的刺。如图 16(a)所示,仙人掌表面长有亲水的锥刺;而宏观尺度上肉眼可见的锥刺,其表面还有着微纳米尺度的锥形倒刺结构。锥形结构可近似看作蜘蛛丝纺锤节结构的一半,也是优异的液滴输运结构单元,可产生 Laplace 压差驱动液滴。
此外,龙舌兰(agaves)、狗尾草(bristlegrass)、麦芒(wheat awns)等植物或植物器官上被发现有类似的锥刺或锥针结构,且具有一定程度的集水能力。如图 16(b)所示,在多尺度的锥刺结构上,小液滴被亲水的倒刺捕捉并由于 Laplace 压差驱动作用而沿着分支输运到大尺度锥刺主道,基于相同原理,主道上的液滴可从大锥刺尖端输运到大锥刺根部。其中,类似水稻叶表面的具有取向的微纳米沟槽结构也起到了引导液滴输运的作用;并且,与蜘蛛丝结构同理,在锥形结构表面的微纳米沟槽所形成的粗糙度梯度,有助于液滴输运。
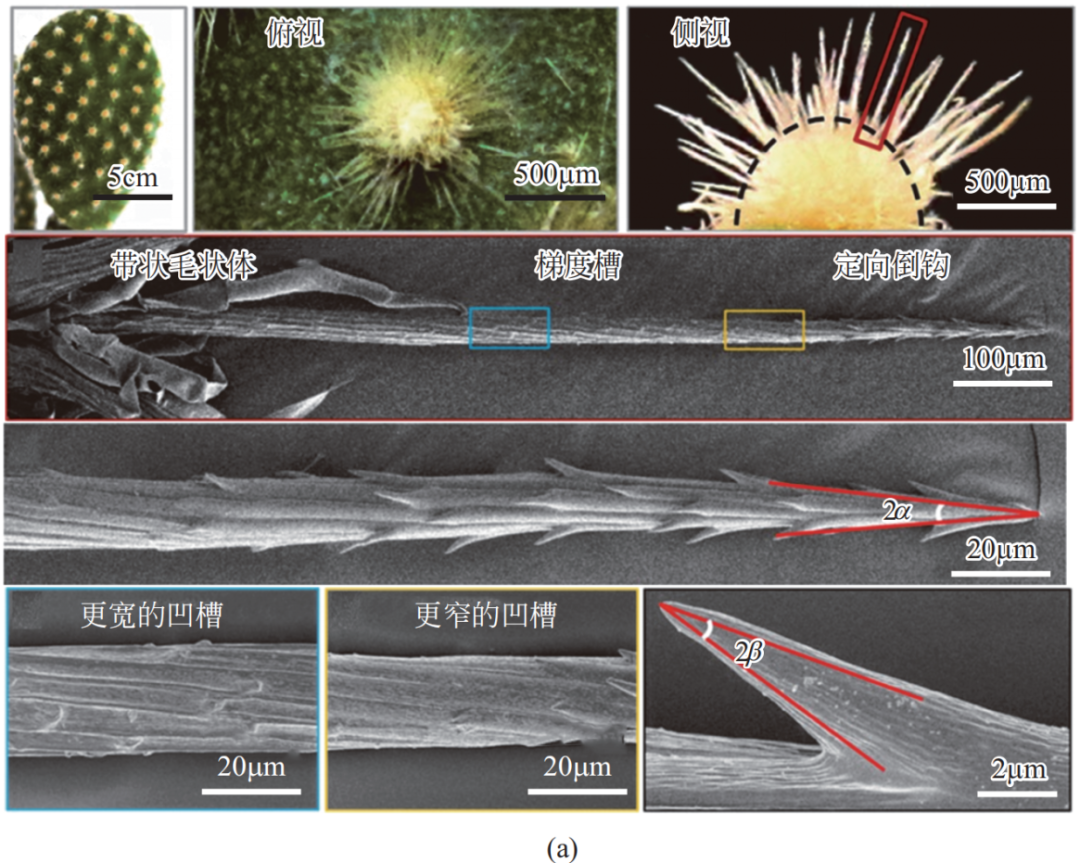

图 16 仙人掌的表面形貌和液滴定向输运
2021 年,Feng 等发现南洋杉叶由锯齿结构组成,每片锯齿状小叶片具有双重曲率结构,包括横向曲率和纵向曲率,如图 17(a)~(c)所示。研究发现,该复杂结构可使乙醇沿锯齿结构取向流动[图 17(d)],而水则沿逆锯齿方向流动[图 17(e)]。如图 17(f)所示,不同表面张力的流体(即不同比例的乙醇 - 水混合物)在南洋杉叶上流动时可产生不同方向和大小的位移。该发现为“不改变表面结构、表面化学组成,不借助外力或能量输入,仅通过调控液体性质实现操控液滴输运”的策略提供了全新的仿生依据。
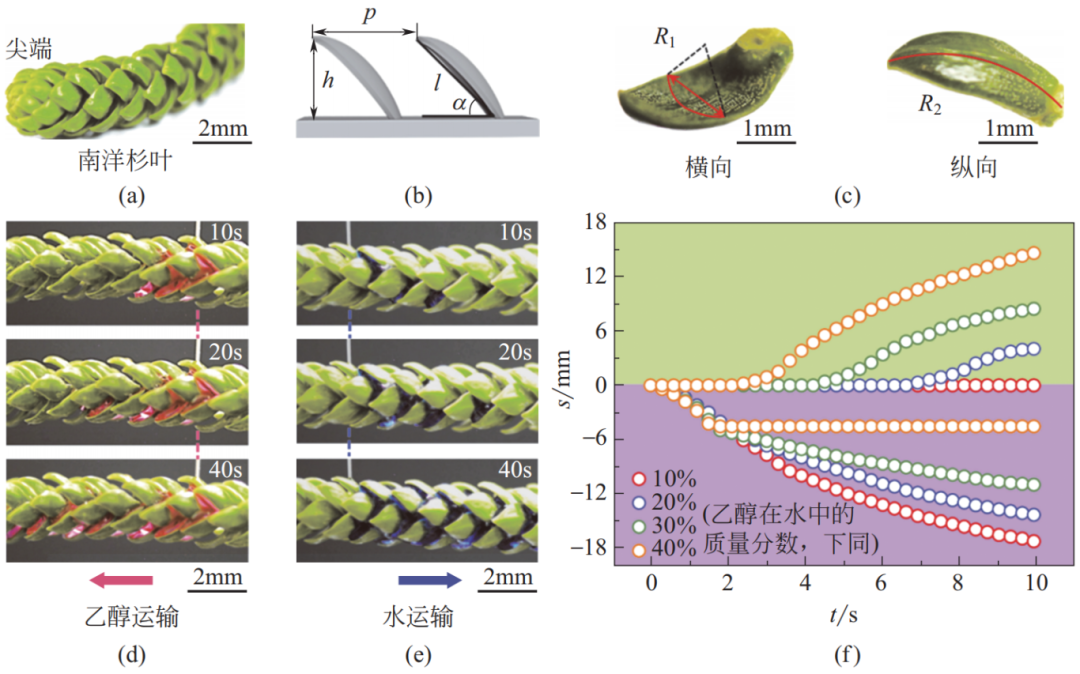
图 17 南洋杉叶的表面形貌及不同液滴在其上不同的输运方向
超亲水表面上液滴与水的黏附力达到理论最大值,液滴虽难以呈球形滚动并脱离表面,但可在表面铺展,在表面上的运动形式可表现为水流。而在自然界,动植物利用超亲水界面使水扩散、流动以适应生存环境,也属于一种操控液滴行为的方式。理论上来说,超亲水要求材料具有很高的表面能以克服表面张力,使液滴自发铺展。
有关超亲水生物表面的研究进展与前文提及的另一极端——超疏油表面情况类似,由于条件苛刻,在自然界尤其是空气环境中发现的实例不多。
很久以前,生物领域科学家发现,人类的眼角膜(cornea)前的泪膜(precorneal tear film)可以使眼泪迅速散开,是典型的超亲水生物表面,其不仅可润湿眼球,还可避免光的散射。如图 18 所示,角膜前的泪膜自内向外可分为黏液层、水样层和脂质层三部分。其中,黏液层一般厚度在0.02 ~ 0.05mm,主要成分为黏蛋白,与角膜结构连接;水样层是泪膜的主体,厚度一般在 6 ~ 10mm,由水、水溶性物质/亲水物质组成,使泪膜具有超亲水及相关功能;脂质层厚度在 0.05 ~ 0.5mm,主要作用是保护水样层,避免水样层与外界直接接触。
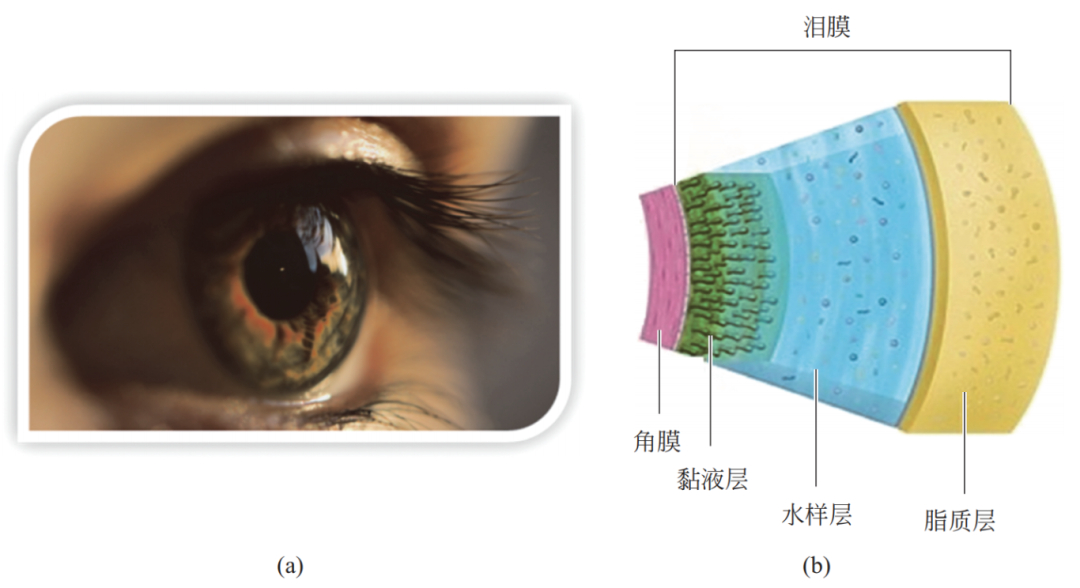
图 18 人类的角膜前泪膜结构及超亲水性
此外,前文提及了近年来研究发现红瓶猪笼草叶笼的唇口具有超亲水性。而红瓶猪笼草叶笼的口袋(pitcher)中,超疏水低黏附的红色“滑移区”相连的下半部分为偏绿色的“消化区(digestive zone)”,该区域由亲水性的生物化学物质构成,表现出高度亲水乃至超亲水性,这有利于水分、消化液的扩散,以淹死或杀死昆虫,分解虫体营养物质。自然界其他的超亲水实例主要是一些生活在水中的生物,表现为水下超疏油低油黏附性质。
超亲水表面在水下环境中可表现为水下超疏油低油黏附的界面性质,本小节主要介绍相关水生生物。对自然界水下超疏油界面深入研究后发现,亲水化学成分和微纳米复合粗糙表面结构是构成水下超疏油界面的关键。
研究表明,荷叶“出淤泥而不染”不仅因为其上表面的超疏水性、自清洁效果,还与其下表面具有的水下超疏油性[水下油接触角(OCA)为 155.0°±1.5°,1,2-二氯乙烷]以及抗油污黏附功能[水下油滚动角(OSA)为 12.1°±2.4°,1,2-二氯乙烷]有关。如图 19 所示,荷叶下表面具有层次微纳米粗糙结构,由长 30 ~ 50μm、宽 10 ~ 30μm、高 4μm的扁平状突起组成,突起表面覆盖有大小为 200 ~ 500nm 的纳米凹槽。

图 19 荷叶下表面的形貌及水下超疏油低油黏附性质
此外,荷叶下表面没有疏水蜡质,而仅由亲水物质组成,使得水分子可嵌入粗糙结构内部,防止油滴浸润,从而实现较稳定的水下超疏油低油黏附特性。
马面鱼(绿鳍马面鲀,Navodon Septentrionalis;鲀鱼,filefish)在被油污染的水域中也可自由游动,水中的油滴会从头到尾定向滑过其表皮而不黏附。研究表明,这主要是由于该种鲀鱼的表皮不仅包含大量的胶原蛋白等高表面能有机物,还长有具有特定取向的钩状棘刺,如图 20 所示。

图 20 鲀鱼表皮的形貌及水下超疏油定向自清洁功能
H—头;T—尾;H →T—从头到尾
这层紧密排列的定向钩状棘刺会增加鱼皮的表面粗糙度,有助于保持鱼皮的水下超疏油低黏附性质。此外,钩状棘刺可增强油滴单向运动趋势,防止油聚集在鱼的头部,赋予鱼在油污水中定向自清洁的能力。
蛤蜊内壳包含两个不同浸润性的区域:无法抗油滴黏附的边缘区域和具有水下超疏油性的中央区域。研究发现,这两个区域的主要化学组成类似,均为亲水的无机碳酸钙;但两者表面结构却存在巨大差异。
如图 21 所示,外围区域的片状碳酸钙整齐排列,表面较平整光滑;内部区域的块状碳酸钙无序堆积,具有很高的粗糙度。亲水性的碳酸钙组成使蛤蜊壳在水中具有疏油性,蛤蜊壳内部区域粗糙微纳米结构有利于将水分子锁在内部表面,降低蛤蜊壳与油滴的接触面积,保障其水下超疏油低油黏附的浸润特性。
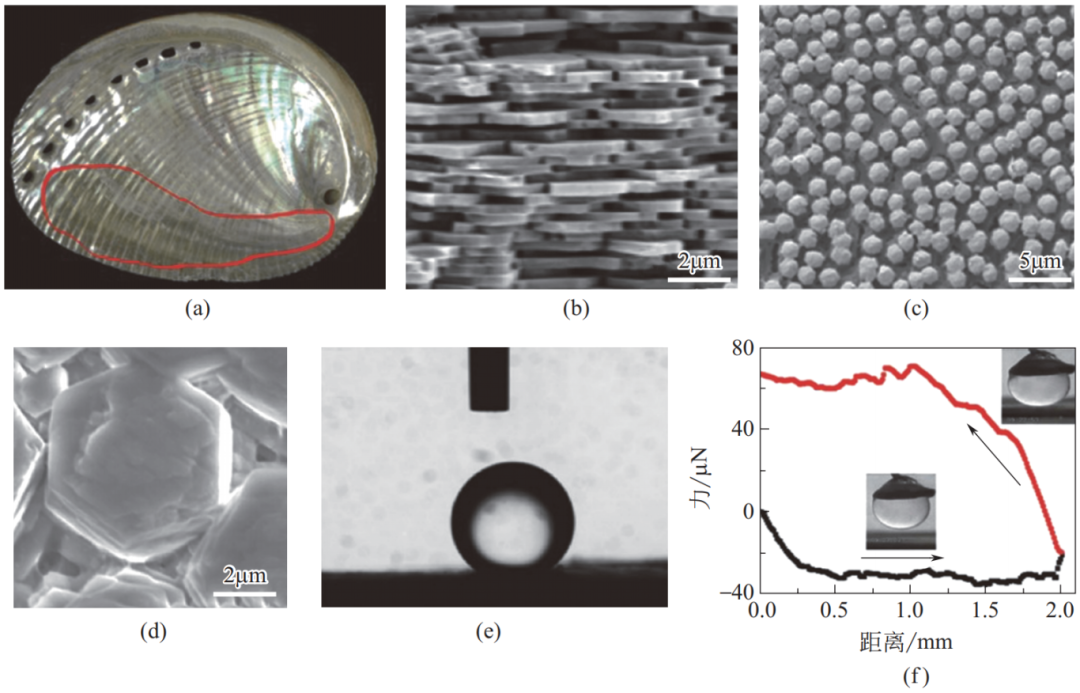
图 21 蛤蜊内壳的表面形貌及水下超疏油低油黏附性质
鱼鳞一度以其微纳米结构的水下减阻功能闻名,除此之外,研究人员发现鱼鳞还具有水下抗油滴浸润能力,这可以让鱼在游经含油水域时不受油污影响。其奥秘在于鱼鳞的化学组成及微纳米结构。鱼鳞的主要成分为亲水性的磷酸钙,且最外层附有一层薄薄的蛋白质黏液层;每个鱼鳞片的表面都分布着许多长 100 ~ 300μm、宽 30 ~ 40μm 的取向性突起,该突起结构具有纳米级的粗糙度,如图22所示。这种化学组成和微纳米结构可将水分子嵌入其中,并形成拒油水垫以抗油污黏附,从而赋予鱼鳞表面优异的水下疏油性能。

图 22 鱼鳞的表面形貌及水下抗油滴浸润 / 黏附性质
Zhang 等研究虾的外壳发现,其表面具有均方根(root mean square,RMS)为 8nm 的粗糙结构,如图 23 所示。该结构有利于水分子在其表面黏附形成水层,从而使虾壳表面对油滴产生很强的排斥作用。此外,虾壳的主要化学成分为亲水性甲壳素,使得水分子可以在表面被截留,从而保障水层的稳定性,阻止油滴进入。两种效应结合,使虾壳在海水环境中对油滴的黏附力也可较稳定地维持在小于 1μN 的状态(对油滴表现出极低的黏附性)。

图 23 虾壳的表面形貌图及水下超疏油低油黏附性
海藻(saccharina japonica;seaweed)以其特殊的生物化学组成及医药学功用而闻名,其特殊组成也赋予其超浸润性。研究表明,即使在高盐度和高离子强度溶液中,海藻仍表现出持久的水下超疏油性以及对油滴的低黏附性。如图 24 所示,在不同浓度的 NaCl 溶液中(从纯水到完全饱和溶液),界面始终显示出大于 150° 的水下 OCA 和非常低的水下 OSA。海藻表面上存在大量的多孔粗糙结构,并含有大量藻酸盐、角叉菜胶和琼脂等天然多糖。这些多糖分子十分亲水,在高盐度溶液中也极易与水分子键合,使海藻表面形成水层。多孔粗糙微纳米结构和对盐不敏感的多糖分子的共同作用使得海藻表面具有耐盐的水下超疏油低油黏附性。
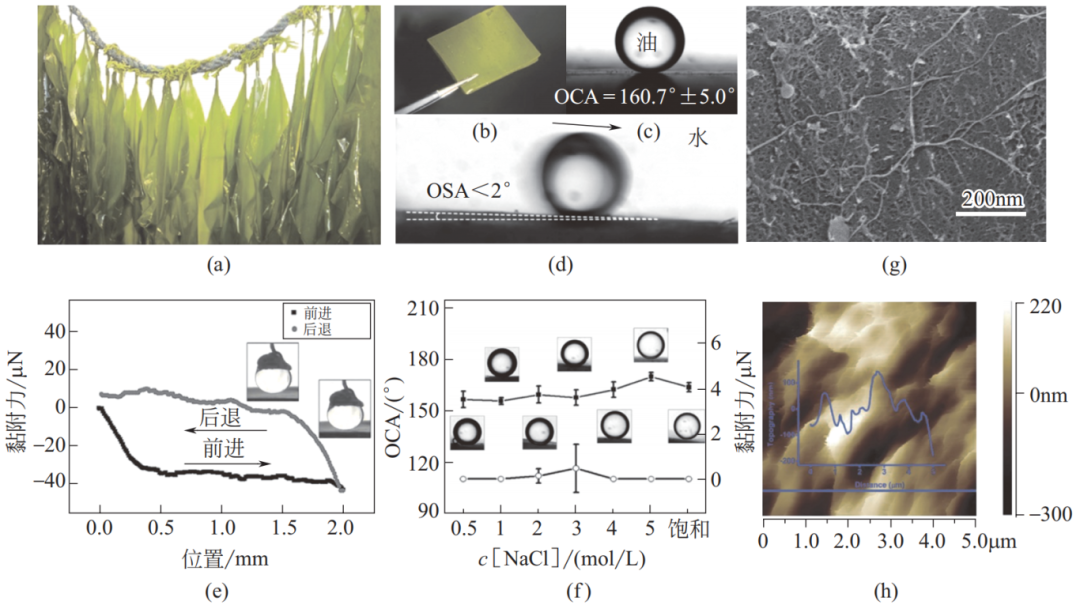
图 24 海藻及其耐盐的水下超疏油低油黏附性
自然界奇妙的生物现象及其背后的科学奥秘随着不断地被深入研究,变得更加鲜活而生动,细致而严谨。除了本次提到的自然界超浸润界面外,还有许多其他仿生超浸润界面,它们在带给研究者以科学启示的同时,也为各类生物技术、仿生材料的实现和应用提供了更多的构筑策略和可能性,相关内容将在后续章节展开介绍。自然之美和科学之美的精妙融合,让仿生超浸润界面未来可期。